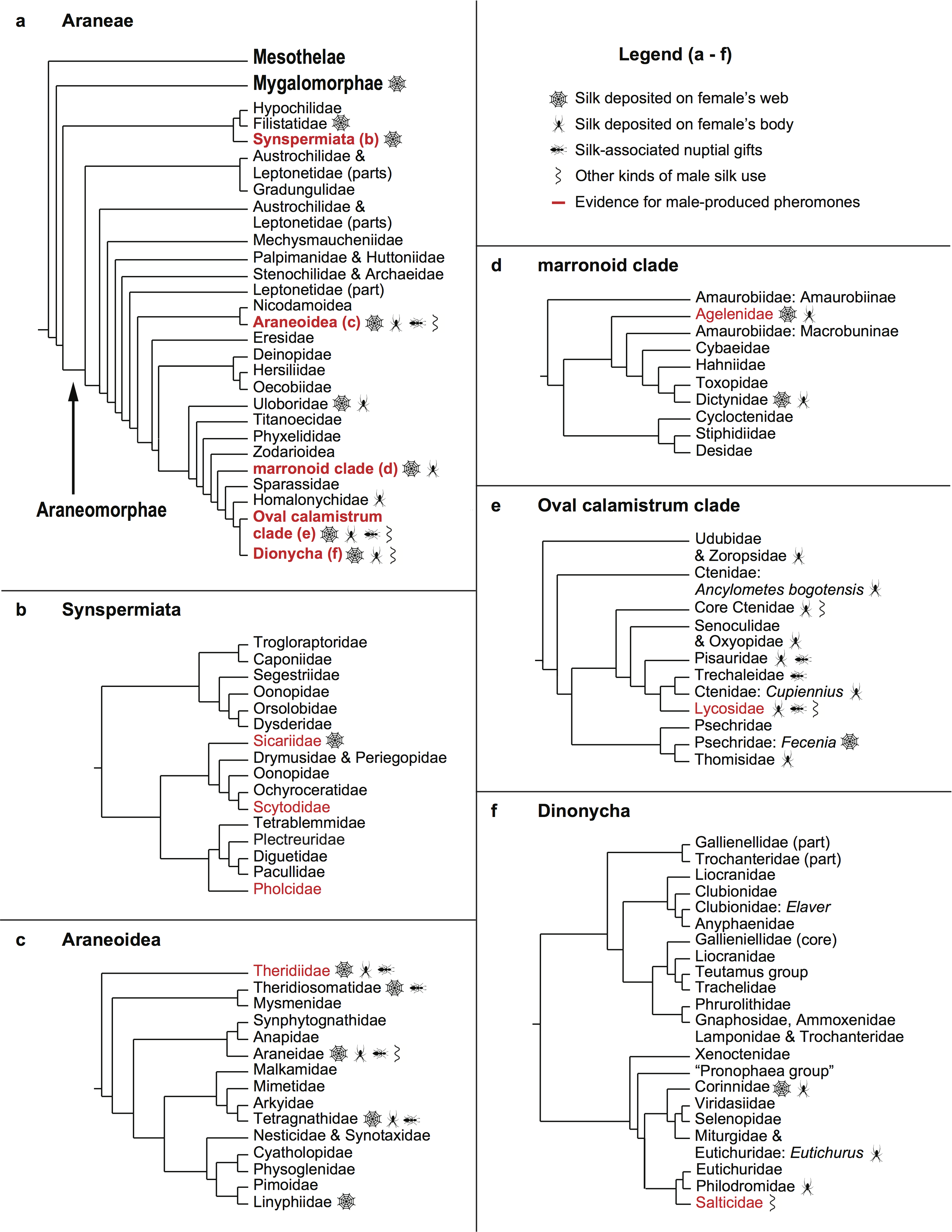
I am very excited to share the publication of a new paper coauthored with Sean McCann and my supervisor Maydianne Andrade in the journal Proceedings of the Royal Society B. The full paper can be found here (please email me or contact me on twitter if you don’t have access and would like a pdf copy). Before I summarize the study below, I would like to first thank the Tsawout First Nation for allowing me and my collaborators to study black widows on their beautiful lands for the last several years. We also offer heartfelt thanks to the many generous contributors* to our #TeamBlackWidow crowdfunding campaign, without whom this research would not have possible. Finally, we are also very grateful to the Toronto Entomologists’ Association for funding part of the fieldwork with the Eberlie Grant, and to NSERC for funding our lab’s research and my PhD program.

Female (left) and male (right) western black widow spiders (Latrodectus hesperus). Photo: Sean McCann
For solitary animals that reproduce sexually, finding a partner is a critical first step in the sequence of events that lead to mating. We know a lot about the traits that help males win fights over females (like horns and other weapons), as well how female preferences can lead to the evolution of extravagant ornaments and displays like the tails of male peacocks (or peacock spiders). But these two mechanisms of sexual selection (male competition and female choice) can often only operate on males that actually find females to compete over. In a lot of animals, including many terrestrial arthropods like insects and spiders, a race to find females determines which males get the opportunity to pass on their genes. The kinds of traits that help males to win this race are less well understood than male ornaments and weapons, in part because it can be tricky to track mate searching males in nature.

Our field site on the sand dunes of southern Vancouver Island, British Columbia. At this site, driftwood logs provide shelters for western black widows. Photo: Sean McCann
In this study, my coauthors and I used experiments and observations of the natural movements made by black widow males to learn more about what gives them an edge when it comes to finding females. First, we spent about six months in the field tracking hundreds of male black widows in their natural habitat. We marked all of the females (who generally stay put on their capture webs, which makes them relatively easy to keep track of) and males (who actively search for females) that we encountered during the season. This allowed us to estimate how many males survive the trip to find females (only about 12%!) and how far they move when they do survive (in most cases, less than 60 metres, but sometimes more than 200, which is pretty impressive for spiders with a body length of less than 1 cm!). We were also able to determine that males outnumber receptive females by more than 10 to 1 during the height of the mating season, which means that competition over mates is fierce, making traits and tactics that confer an advantage to searching males all the more important.

We spent a lot of nights on the beach checking the webs of marked females, marking and recording any males who visited them. Photo: Sean McCann
Before we get to those tactics, however, let me back up for a minute and summarize some important features of black widow sexual communication and behaviour that are relevant to this story. First, female black widows produce a sex pheromone that functions as a chemical personal ad. This chemical message is released from the silk of a female’s web, and it provides males with information about her location and sexual receptivity. Before this study, we didn’t know the range of this message, just that it operates over some distance, allowing males to locate females who are ready to mate. Male black widows detect the female’s pheromone using sensory hairs on their legs. Once a male finds a female, he engages in a courtship dance that transmits vibrations through her web, providing her with information about his identity and quality as a mate. After several hours of dancing and laying down silk all over the web and the female’s body (this silk may contain chemical messages just like the female’s), and assuming he is not interrupted by a rival or eaten by the female (it happens, particularly if she’s hungry and therefore more interested in a meal than mating!), the male eventually mates with her. Spiders do this in a strange and unique way, transferring sperm with paired copulatory organs called pedipalps. The first male to mate can break off the tips of his copulatory organs inside the female, effectively blocking rival males from inseminating her, and thus ensuring his paternity.

A female (yellow 567, whose number corresponds to the location of the paint marks on her legs) consumes an unlucky male. Photo: Sean McCann.
Ok, now back to the research! The experimental part of our study involved setting up a series of actual races for male black widows—first longer-distance contests in the field, and then shorter sprints in the laboratory. For the great black widow races of 2016 and 2017, we set up a 60-metre course on the sand dunes at our field site. The finish line was made up of a series of mesh cages containing females and their silk.

The finish line of the 2016 great black widow race. Each cage contains a female black widow on her pheromone-emitting web. Photo: Sean McCann
Before the race, we weighed in each male on a tiny scale**, measured the length of his legs, and painted him with unique racing stripes so we would be able to track whether he completed the race and calculate his average speed. At sunset (black widows are nocturnal, so males search for females at night) we released groups of about 20 males at 10 metre intervals from the finish line, so that the closest group only had to travel 10 metres and the farthest group had to travel 60 metres. The course was set up so that males would be downwind of the line of pheromone-emitting females (assuming that the forecast was correct), and once all of the males were released, we waited at the finish line for them to start arriving outside of females’ cages.

A green-marked male crosses the finish line! Photo: Sean McCann
In 2016, when the wind was strong and came fairly consistently from the forecasted direction, males released at all distances up to 60 metres were equally likely to find females, which suggests that they are very sensitive to the smell of females. In 2017, however, when the wind ended up being weak and highly variable in direction, males released farther than 40 m from females were never able to locate them. Clearly, wind speed and direction will strongly affect the ability of a male to detect and find a female using only their sense of smell. But these experiments also revealed something surprising. In 2016, we found that the males that started out farthest from females achieved the fastest average speeds during searching—up to almost 1.5 metres (or more that 150 body lengths for spiders that are typically less than 1 cm long) per minute! And in 2017 we found that not only were males able to reach the finish line even after the wind shifted so much that smelling females on the racecourse was likely impossible, but also that the vast majority of males ended up outside of the cage of the single female who was in line with the direction of the wind during the first couple hours of the experiment.

A yellow-marked male makes his way down the race course. Photo: Sean McCann
Spending time watching what these spiders actually do when they search for females helped us make sense of these results. Male black widows have very poor vision, so they are guided toward females by their sense of smell. To get their bearings, they climb up vegetation, and wave first pair of legs (which you’ll recall are covered with sensory hairs), apparently ‘tasting’ the wind. After a while, they will climb back town to the ground and continue to move toward their target. Like all spiders, male black widows trail silk draglines behind them as they move through their environment. They anchor these safety lines to the vegetation periodically, such that they leave a silk path wherever they go. We noticed that when a searching male encounters one of these trails, he runs along it, using it like a silk highway. We realized that if males recognize the silk of rival males, they may use their trails to find females, even if wind conditions make it difficult to smell a female directly. And since these spiders are much more adept at walking on silk than on the ground, this might explain why the males in our experiments who traveled farthest also traveled fastest. The males released at 60 metres would have been most likely to encounter the silk trails left behind by all those who were released closer to the females.

A white-marked male has climbed up some grass and is extending his forelegs to taste the air for female pheromones. Photo: Sean McCann
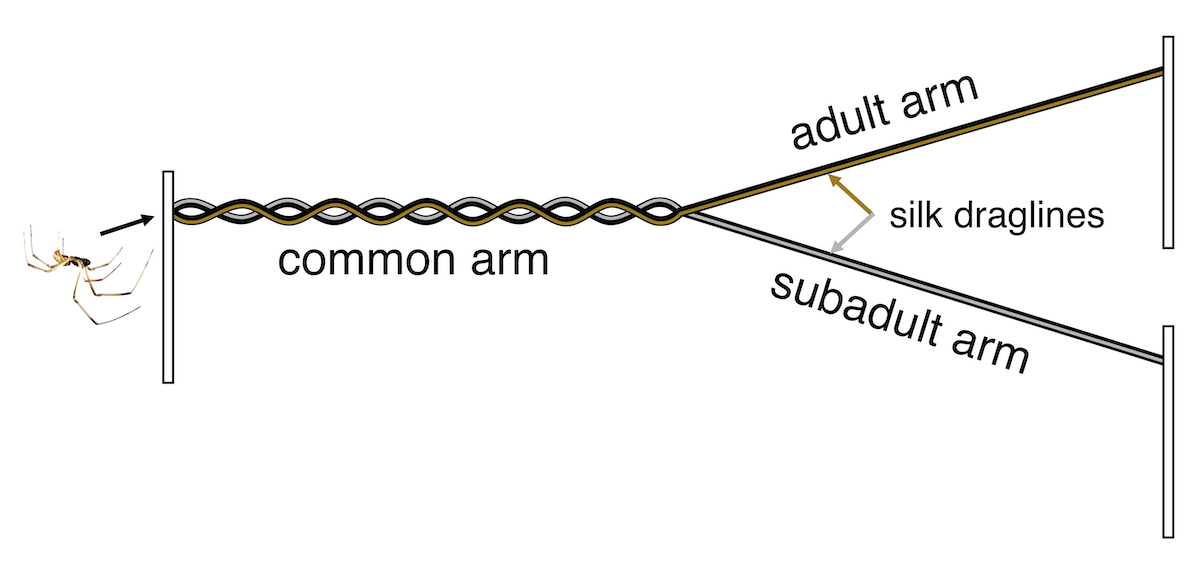
To test this idea directly, we next ran a series of experiments that we called the X-races, the first of which was carried out by students in the behavioural ecology class Sean and I taught together at UTSC in 2017. The race course in this experiment is an X-shaped maze made out of string, with a female set up at one end with a fan behind her to blow her pheromone toward the male, who is released on one of the short arms of the X at the end opposite the female.
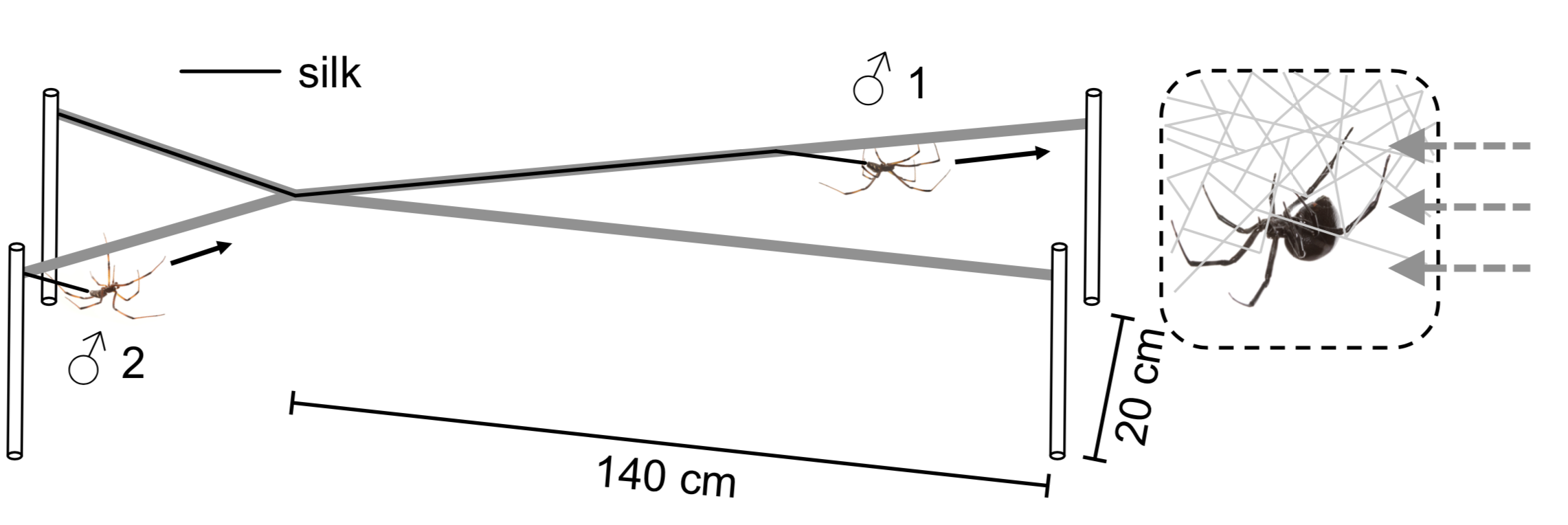
The X-race, an experimental setup for testing whether males follow or avoid the silk of rival males.
After being placed on the string, he moves upwind and when he reaches the centre of the X, he can choose to follow either arm—both lead to the female, and either way, he leaves a trail of silk behind him. Next, we introduce a second male to the end of the maze farthest from the female, but on the opposite arm from the first male. When this male gets to the intersection of the X, he now has a choice to follow or avoid the silk of the rival male. We used this experimental set up to confirm that males follow the silk of rivals when given the choice, and that they travel faster when they do. Using a modified version of the X-race, we also found that males only follow the silk of other black widows (and not that of closely related false widow males, who also occur at our field site), which means that the information on the silk is species-specific.

Sean (foreground) and our 4th year behavioural ecology students set up an X-race in the lab. The trays of water below prevent the spiders from leaving the maze if they drop down on a dragline.
When we put all this together, it tells us that male black widows use the somewhat surprising tactic of following their rivals to find females faster, and that exploiting the silk trails produced by earlier searchers allows them to locate females efficiently even when conditions make it difficult to directly detect their chemical messages. It might not seem like a great idea to follow another male to a female’s web, because this guarantees a competition over the opportunity to mate with that female. We would expect male animals to use cues about the presence of rival males to avoid competition, when given the choice (and in at least one other spider species, they do). But our time following this population in the field revealed that these males are unlikely to ever have that choice. There are so few sexually receptive females signaling on any given night that competition is inevitable. In this situation, the best tactic for males may be to arrive at a female’s web as fast as possible, even if other males are already there. Although being the first male to mate is important for black widows, being first to arrive is not critical, because courtship may last several hours. It’s at this point, at the female’s web, that competition and female choice can finally kick in.

Two males who have arrived at the same female’s web one after the other. Photo: Sean McCann
We are excited about these results, which reveal a surprising means of using indirect information to gain a competitive edge in the race to find females. We are also hope that our experimental race designs can allow us and other researchers to learn about spider mate searching behaviour and chemical communication in the future. Setting up races over different kinds of terrain to look at the effects of physical barriers on male performance, or doing races over several days and longer distances could yield more insights into what traits are important for searching males. Longer distance ultramarathons for spiders might be more appropriate in environments where females are more widely dispersed than at the site we studied. And the X-race is a convenient way to test male decision-making under controlled laboratory conditions while using a setup that reasonably reflects how male spiders actually move in the field.
This is only the first chapter of black widow behaviour research brought to you by the support of #TeamBlackWidow, and we look forward to sharing the next instalment soon!
 Thank you SO MUCH to Catherine & Doug Antone, Joe Lapp, Robb Bennett, Roy Dunn, Sean Lambert, Betty Kipp, Dora Sardas, Kristen Cain, Christy Peterson, Christy Peterson, Raphael Royaute, Dawn Bazely, Woodrow Setzer, Pierre Robillard, John Barthelme, Nemo de Jong, Mike Boers & Tanya Stemberger, Sina Rastegar, Sarah Langer, Sidnee & John Scott, Stephen & Linda Lambert, Staffan Lindgren, Amanda Yee, Rob Higgins, Tonia Harris, Tanya Jones, Joe O’Franklin, Dezene Huber, Tracey Birch, Peggy Muddles, Regine & Gerhard Gries, Gwylim Blackburn & Samantha Vibert, Alex & Karla Antone, Gil Wizen, Gwen Pearson, Joan Andrade, Kate Compton, Peggy McCann, Peter Andrade, Rick Redus, Robyn Raban, Shelley Barkley, Stewart, Geoff Bennett, Kyle Cassidy, Colin & Heather McCann, Jonathan Meiburg, Lori Weidenhammer, Diana Davis, Ray Scanlon, Ashley Bradford, Ed Morris, Robert Cruickshank, Marc Rashinski, James Petruzzi, Joseph Peter McNamara, Ariel Ng, Robert Neylon, Auriel Fournier, Victoria Nations, Leah Ramsay, Tom Pearce, Chloe Gerak, Scott Severs, Angie Macias, Nick Spencer, Thomas Astle, Luna Nicolas Bradford Ley, Peter Midford, Laurel Ramseyer, Morgan Vis, Tom Pardue, Scott Schrage, Kelly Brenner, Karen Yukich, Charmaine Condy, Amy Parachnowitsch, Catherine Scott, Christine Rock, Jason Parker-Burlingham, Jonathan Kade, Joseph Peter McNamara, Joshua Erikson, Juniper English, Nick Spencer, Robert Cruickshank, Sabrina Caine, Suran TheStorm, Richard Dashnau, Stephen Heard, Holly Fraser, Lynne Kelly, Roberta Chan, Kat Cruickshank, Meera Lee Sethi, Mike Hrabar, Tiffany Jacobs, Connie Larochelle, Willow English, David Steen, Michelle Reeve, Tone Killick, David Esopi, Antonia Guidotti, Elaine Wong, Lisa Wrede, Naomi Gonzales, Don Campbell, Matt Masterson, Paul Manning, Casey Peter, Dave Rich, Jessical Olin, Kate Rey, Katie Russell, Shari McDowell, Suzanne Spinelli, Christina Tran, Cindy Wu, Aaron Soley, Chris Garbutt, Greg Randolph, Lila Robinwood, Eric Damon Walters, The Spider Chick, & Steve Waycott!!!
Thank you SO MUCH to Catherine & Doug Antone, Joe Lapp, Robb Bennett, Roy Dunn, Sean Lambert, Betty Kipp, Dora Sardas, Kristen Cain, Christy Peterson, Christy Peterson, Raphael Royaute, Dawn Bazely, Woodrow Setzer, Pierre Robillard, John Barthelme, Nemo de Jong, Mike Boers & Tanya Stemberger, Sina Rastegar, Sarah Langer, Sidnee & John Scott, Stephen & Linda Lambert, Staffan Lindgren, Amanda Yee, Rob Higgins, Tonia Harris, Tanya Jones, Joe O’Franklin, Dezene Huber, Tracey Birch, Peggy Muddles, Regine & Gerhard Gries, Gwylim Blackburn & Samantha Vibert, Alex & Karla Antone, Gil Wizen, Gwen Pearson, Joan Andrade, Kate Compton, Peggy McCann, Peter Andrade, Rick Redus, Robyn Raban, Shelley Barkley, Stewart, Geoff Bennett, Kyle Cassidy, Colin & Heather McCann, Jonathan Meiburg, Lori Weidenhammer, Diana Davis, Ray Scanlon, Ashley Bradford, Ed Morris, Robert Cruickshank, Marc Rashinski, James Petruzzi, Joseph Peter McNamara, Ariel Ng, Robert Neylon, Auriel Fournier, Victoria Nations, Leah Ramsay, Tom Pearce, Chloe Gerak, Scott Severs, Angie Macias, Nick Spencer, Thomas Astle, Luna Nicolas Bradford Ley, Peter Midford, Laurel Ramseyer, Morgan Vis, Tom Pardue, Scott Schrage, Kelly Brenner, Karen Yukich, Charmaine Condy, Amy Parachnowitsch, Catherine Scott, Christine Rock, Jason Parker-Burlingham, Jonathan Kade, Joseph Peter McNamara, Joshua Erikson, Juniper English, Nick Spencer, Robert Cruickshank, Sabrina Caine, Suran TheStorm, Richard Dashnau, Stephen Heard, Holly Fraser, Lynne Kelly, Roberta Chan, Kat Cruickshank, Meera Lee Sethi, Mike Hrabar, Tiffany Jacobs, Connie Larochelle, Willow English, David Steen, Michelle Reeve, Tone Killick, David Esopi, Antonia Guidotti, Elaine Wong, Lisa Wrede, Naomi Gonzales, Don Campbell, Matt Masterson, Paul Manning, Casey Peter, Dave Rich, Jessical Olin, Kate Rey, Katie Russell, Shari McDowell, Suzanne Spinelli, Christina Tran, Cindy Wu, Aaron Soley, Chris Garbutt, Greg Randolph, Lila Robinwood, Eric Damon Walters, The Spider Chick, & Steve Waycott!!!
^Thanks very much to Jay Cullen at UVic for kindly allowing us and our spiders access to his lab and microbalance!